| |||||||||||||||||||||||||||||||||||||||
| Enter search term: | ||||
| International Journal of Antimicrobial Agents | ||
| Vol: 22 Issue: 2, August, 2003 | Article Full Text PDF (208 KB) | |
| pp: 155 - 159 | Bibliographic Page | |
| Full Text Article HTML | ||
| Get citation export (Reference Format) | ||
The major lysosomal cysteine proteinase of African trypanosomes is a candidate target for novel chemotherapy of sleeping sickness. This cathepsin L-like enzyme is termed rhodesain and brucipain in Trypanosoma brucei rhodesiense and Trypanosoma brucei brucei, respectively. Three potent and selective dipeptidyl cathepsin L inhibitors have been investigated for their trypanocidal activities in vitro using culture-adapted bloodstream forms of T. b. brucei. Compared with general cysteine proteinase inhibitors used previously by ourselves and others, the present inhibitors had improved selectivity indices and, importantly, anti-trypanosomal activities comparable with those of commercial anti-sleeping sickness drugs. Using purified recombinant rhodesain, potent kinact/ Ki values of up to 2.3×106 M-1 s-1 were recorded with the inhibitors. Also, all inhibitors blocked proteinolysis in the lysosome consistent with the inhibition of rhodesain/brucipain. In conclusion, the data support the potential of cathepsin L inhibitors for rational anti-trypanosomal drug development.
African trypanosomes, the causative agents of sleeping sickness in humans and nagana disease in cattle, are flagellated protozoa that live extracellularly in the blood and tissue fluids of their mammalian host and are transmitted by the bite of infected tsetse flies. Over 60 million people living in 36 countries are at risk of acquiring sleeping sickness and the estimated number of people thought to have the disease is between 300 000 and 500 000 [ 1 ]. In addition, 46 million cattle are exposed to the risk of contracting nagana and the disease costs an estimated US $1340 million per year [ 2 ]. Chemotherapy of African trypanosomiasis still relies on drugs developed decades ago and some of these show toxic side effects [ 3 ]. In addition, the emergence of drug-resistant trypanosome strains has been reported [ 4, 5 ]. Thus, new strategies to treat African trypanosomiasis are required.
One promising line of research towards the establishment of new anti-trypanosomal chemotherapies has been the targeting of the major cysteine proteinase of the parasite [ 6, 7, 8 ]. In African trypanosomes, the major lysosomal cysteine proteinase has a primary sequence and biochemical characteristics broadly similar to mammalian cathepsin L [ 9, 10, 11 ]. The enzyme is termed rhodesain and brucipain in Trypanosoma brucei rhodesiense and Trypanosoma brucei brucei, respectively. Recently, irreversible, dipeptidyl, substrate-analogue inhibitors of cysteine proteinases were shown to kill T. b. brucei, the model organism for human African trypanosomes, both in culture and experimentally infected animals [ 7, 12, 13 ]. Importantly, it was demonstrated that killing of trypanosomes was correlated with inhibition of the target enzyme brucipain [ 12 ]. However, so far, only dipeptidyl inhibitors with a broad activity against cysteine proteinases have been tested. For this report, therefore, we have tested the trypanocidal activities and inhibition efficacies of three commercially available cathepsin L inhibitors.
N-Benzyloxycarbonyl-phenylalanyl-arginine-7-amido-4-methylcoumarin (Z-Phe-Arg-AMC), N-benzyloxycarbonyl-phenylalanyl-alanyl diazomethylketone (Z-Phe-Ala-CHN2), N-benzyloxycarbonyl-phenylalanyl-tryrosinal (Z-Phe-Tyr-CHO), N-benzyloxycarbonyl-phenylalanyl- t-butyl-tryrosyl diazomethylketone (Z-Phe-Tyr(OtBu)-CHN2), and 1-naphthalenylsulphonyl-isoleucyl-tryptophanal (1-NapSO2-Ile-Trp-CHO) were from Bachem (Heidelberg, Germany or Torrance, CA, USA). 1,4-Diazabicyclo[2.2.2]octane (DABCO), 4,6-diamidino-2-phenylindole (DAPI), and bovine serum albumin (BSA) were from Sigma (Dorset, U.K.). FITC-labelled human transferrin was from Cambridge BioScience (Cambridge, UK).
The substrate Z-Phe-Arg-AMC and inhibitors were prepared as 10 and 20 mM
stock solutions, respectively, in DMSO. Assays were performed at room
temperature and incorporated either recombinant rhodesain [ 8 ], or, as a comparison, recombinant cruzain [ 14 ] derived from the South American trypanosome, T. cruzi.
Rhodesain (8.3 nM) or cruzain (7.5 nM) was pre-incubated for 20 min at room
temperature in 100 ml of 100 mM sodium acetate, pH 5.5, 5 mM DTT, in the presence of
inhibitor (0.00160 mM) or DMSO alone added as a 1
ml aliquot. For rhodesain, the incubation buffer also
contained 0.01% Triton X-100. Substrate (10 mM), in 100
ml of the appropriate buffer, was then added. Release
of free AMC was measured at emission and excitation wavelengths of 355 and 460
nm, respectively, in a Molecular Devices Flex-Station fluorometer. Measurements
were taken every 1.52 s. The rate of inhibition ( kobs) was
determined at each inhibitor concentration according to [ P]=
vo/ kobs×{1-exp(- kobs×
t)}, where [ P] is the concentration of product formed over time
t and vo the initial rate, using non-linear regression
analysis ( 
 3.0a). If the kobs
versus inhibitor concentration could reliably fit a two-step irreversible
mechanism ( r2>0.9), the inhibition constant
Ki and the inactivation constant kinact were
determined by non-linear regression analysis according to
kobs=
kinact×[I]o/{[I]o+
Ki*×(1+[S]o/ Km)} where
[I]o and [S]o are the concentrations of inhibitor and
substrate, respectively, and Ki*=
Ki×(1+[S]o/ Km). For those
inhibitors with which kobs was linear with increasing
concentrations of inhibitor, linear regression analysis was used to obtain the
association constant kass using kobs=
kass×[I]o/(1+[S]o/
Km). Multiple determinations at each inhibitor concentration
were performed.
3.0a). If the kobs
versus inhibitor concentration could reliably fit a two-step irreversible
mechanism ( r2>0.9), the inhibition constant
Ki and the inactivation constant kinact were
determined by non-linear regression analysis according to
kobs=
kinact×[I]o/{[I]o+
Ki*×(1+[S]o/ Km)} where
[I]o and [S]o are the concentrations of inhibitor and
substrate, respectively, and Ki*=
Ki×(1+[S]o/ Km). For those
inhibitors with which kobs was linear with increasing
concentrations of inhibitor, linear regression analysis was used to obtain the
association constant kass using kobs=
kass×[I]o/(1+[S]o/
Km). Multiple determinations at each inhibitor concentration
were performed.
Bloodstream forms of the monomorphic T. b. brucei clone 427-221a [ 15 ] were grown in Baltz medium [ 16 ] supplemented with 16.7% (v/v) heat-inactivated foetal bovine serum. Human HL-60 cells (DSMZ, Braunschweig, Germany) were propagated in RPMI 1640 medium supplemented with 10% (v/v) heat-inactivated foetal bovine serum. All cell cultures were maintained in a humidified atmosphere containing 5% CO2 at 37 °C.
Toxicity studies were performed as previously described [ 12 ]. Cells were seeded into 24-well tissue culture plates in 1 ml medium containing various concentrations (10-410-12 M) of cathepsin L inhibitors dissolved in 100% DMSO. The controls contained DMSO alone. In all experiments, the final DMSO concentration was 1%, which had no effect on cell growth [ 17 ]. To ensure that the cells were in logarithmic growth phase during the entire experiment, they were seeded at an initial density of 1×104 trypanosomes ml-1 and 5×104 HL-60 cells ml-1, respectively. After 48 h of incubation, the cells were counted using a Neubauer haemocytometer. Each experiment was repeated three times. The 50% effective dose value (ED50), i.e. the inhibitor concentration necessary to reduce the growth rate of the cells to 50% of that of controls, was determined by linear interpolation as previously described [ 18 ]. The minimum inhibitory concentration (MIC), i.e. the lowest concentration of an inhibitor at which all cells were killed, was determined microscopically.
Bloodstream forms of T. b. brucei 427-221a (5×106 per ml)
were incubated with 100 mg ml-1
FITC-labelled human transferrin in Baltz medium supplemented with 2% BSA in the
presence or absence of cathepsin L inhibitors. After 2 h, cells were fixed with
2% formaldehyde/0.05% glutaraldehyde in PBS, applied to poly- 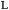 -lysine coated microscope slides, and
treated with 0.0001% DAPI in PBS. The slides were mounted in Mowiol 4-88
[ 19 ] containing 25 mg ml-1 DABCO as anti-fade reagent.
Cells were examined with a Leica DMR microscope using a PL Fluotar 100× oil
objective. Images were recorded using a CCD camera.
-lysine coated microscope slides, and
treated with 0.0001% DAPI in PBS. The slides were mounted in Mowiol 4-88
[ 19 ] containing 25 mg ml-1 DABCO as anti-fade reagent.
Cells were examined with a Leica DMR microscope using a PL Fluotar 100× oil
objective. Images were recorded using a CCD camera.
Three potent cathepsin L inhibitors, Z-Phe-Tyr-CHO [ 20 ], Z-Phe-Tyr(OtBu)-CHN2[ 21 ], and 1-NapSO2-Ile-Trp-CHO [ 22 ] were tested for their anti-trypanosomal activities. All three compounds showed a dose-dependent effect on the proliferation of bloodstream-form trypanosomes with ED50 values in the mid nanomolar range and MIC values between 10 and 100 mM ( - Table: [ 1]). In contrast, the cytotoxicities of the inhibitors towards mammalian HL-60 cells were much lower with ED50 values in the mid micromolar range and MIC values >100 mM ( - Table: [ 1]). As a result, the selectivity index (SI), i.e. the ED50 value obtained with HL-60 cells divided by that found for T. b. brucei, was >100 for all inhibitors ( - Table: [ 1]). The most active compound was 1-NapSO2-Ile-Trp-CHO with an SI of 1150. Compared with previously tested dipeptidyl cysteine proteinase inhibitors [ 7, 12, 13 ], the present cathepsin L inhibitors exhibit at least a 10-fold higher trypanocidal activity against in vitro-cultured T. b. brucei bloodstream forms with respect to ED50 values.
| |||||
Inhibition kinetics of the putative target rhodesain were measured with the three cathepsin L inhibitors and the general cysteine proteinase inhibitor, Z-Phe-Ala-CHN2, one of the most trypanocidal compounds previously tested [ 7, 12 ]. Data were compared with those obtained for cruzain, the related cysteine proteinase of T. cruzi. All three cathepsin L inhibitors were potent inhibitors of rhodesain and cruzain with inactivation rates ranging from 1.5×104 to 4.3×106 M-1 s-1 ( - Table: [ 2]). Two of the cathepsin L inhibitors, namely Z-Phe-Tyr-CHO and 1-NapSO2-Ile-Trp-CHO, yielded better inactivation rates (42- and 11-fold, respectively, with rhodesain) than Z-Phe-Ala-CHN2. For both enzymes, each inhibitor was similarly potent and the same rank order of inhibition was found.
| |||||
Transferrin has been shown to be a lysosomal marker for bloodstream-form trypanosomes [ 23, 24, 25 ]. To determine whether the three cathepsin L inhibitors block proteinolysis in lysosomes, T. b. brucei bloodstream forms were incubated with FITC-labelled human transferrin. In the presence of 100 mM Z-Phe-Tyr-CHO, 10 mM Z-Phe-Tyr(OtBu)-CHN2, or 10 mM 1-NapSO2-Ile-Trp-CHO, the respective concentrations of the inhibitors at which all parasites were killed in vitro (see - Table: [ 1]), a fluorescent spot was observed between the kinetoplast and the nucleus (Fig. 1). No labelling was detected in the absence of inhibitor (Fig. 1). The labelled region corresponded to the location of the lysosome in bloodstream-form trypanosomes [ 23, 24, 25 ]. This result demonstrates that the presence of cathepsin L inhibitors in trypanosome culture leads to accumulation of undegraded transferrin in the lysosome most likely through inhibition of brucipain activity.
| |||||
The major proteinolytic activity in the lysosome of bloodstream-form trypanosomes derives from a cathepsin L-like cysteine proteinase termed brucipain in T. brucei[ 9, 11, 26 ] or rhodesain in T. b. rhodesiense[ 8 ]. Previous studies have shown that dipeptidyl cysteine proteinase inhibitors exhibiting ED50 values in the low to mid micromolar range kill T. b. brucei bloodstream forms in vitro [ 7, 12, 13 ]. Also, only cysteine proteinase inhibitors having alanine, phenylalanine, homophenylalanine or valine at the P1 position (in the nomenclature of Schechter and Berger [ 27 ]) have been investigated for their anti-trypanosomal activities so far [ 13 ]. On the other hand, it has been shown that dipeptidyl compounds with tyrosine or tryptophan in the P1 position preferentially inhibit cathepsin L over other cysteine proteinases [ 20, 21, 22 ]. The results of this study seem to support this observation. Z-Phe-Tyr-CHO, Z-Phe-Tyr(OtBu)-CHN2 and 1-NapSO2-Ile-Trp-CHO, three dipeptidyl compounds selected for potent inhibitory activity against cathepsin L [ 20, 21, 22 ] are, on average, 10100 fold more potent than inhibitors previously tested [ 7, 12, 13 ]. Their ED50 values of 18126 nM compare well with those of the commercial anti-trypanosomal drugs suramin (130360 nM [ 17 ]) and diminazene aceturate (20100 nM [ 17 ]). Though the MIC values of the present inhibitors (10100 mM) are higher than those of suramin (1.42.3 mM [ 17 ]) and diminazene aceturate (0.50.92 mM [ 17 ]), the SI values of 1411150 approach those of the same commercial drugs (1900 for suramin and 3600 for diminazene aceturate [ 17 ]).
A perfect correlation between anti-trypanosomal activity and inactivation rates of rhodesain was not observed for the cathepsin L inhibitors. However, the data indicate that the peptidyl aldehydes are more potent against both the target enzyme and parasites in culture. Discrepancies between enzyme inhibition and trypanocidal activity may be ascribed to differences in the uptake of the inhibitors by the parasite. Interestingly, 1-NapSO2-Ile-Trp-CHO has as N-terminal substituent (P3 position) a naphthyl group which recently has been suggested to facilitate the uptake of respective acyl hydrazide proteinase inhibitors by bloodstream forms of T. b. brucei[ 28 ].
Treatment of T. b. brucei bloodstream forms in vitro with the cathepsin L inhibitors elicited an accumulation in the lysosome of undigested transferrin, which is an established lysosomal marker of African trypanosomes [ 23, 24, 25 ]. Given that brucipain is the predominant cysteine proteinase in T. b. brucei and is localised to the lysosome [ 8 ], it is reasonable to consider that brucipain is the critical target of the cathepsin L inhibitors.
In conclusion, the results suggest that inhibitors of cathepsin L are a rational choice for future anti-trypanosomal drug development. For instance, it would be interesting to test the three compounds coupled to the reactive vinyl sulphone group rather than to the aldehyde or diazomethylketone groups. Vinyl sulphones are relatively novel inhibitors [ 29, 30 ] and are considerable less toxic than aldehyde or diazomethylketones. For example, vinyl sulphone inhibitors have previously been shown to kill bloodstream forms of T. b. brucei in vivo [ 7 ] and to cure experimental infections of mice with T. cruzi without undue toxicity to the host [ 31 ].
| [1]. | WHO. African trypanosomiasis or sleeping sickness. World Health Org. Fact Sheet 2001;259 ( http://www.who.int/inf-fs/en/fact259.html ). |
| [2]. | Kristjanson P.M., Swallow B.M., Rowlands G.J., Kruska R.L., de Leeuw P.N., "Measuring the costs of African animal trypanosomosis, the potential benefits of control and returns to research", Agric. Syst., Volume: 59, (1999), pp. 79-98 Bibliographic Page Full text |
| [3]. | Croft S.L., "The current status of antiparasitic chemotherapy", Parasitology, Volume: 114, Issue: Suppl. (1997), pp. S3-S15 |
| [4]. | Geerts S., Holmes P.H., Diall O., Eisler M.C., "African bovine trypanosomiasis: the problem of drug resistance", Trends Parasitol., Volume: 17, (2001), pp. 25-28 Bibliographic Page Full text |
| [5]. | Matovu E., Seebeck T., Enyaru J.C.K., Kaminsky R., "Drug resistance in Trypanosoma brucei ssp., the causative agents of sleeping sickness in man and nagana in cattle", Microbes Infect., Volume: 3, (2001), pp. 763-770 Bibliographic Page Full text |
| [6]. | McKerrow J.H., Engel J.C., Caffrey C.R., "Cysteine protease inhibitors as chemotherapy for parasitic infections", Bioorg. Med. Chem., Volume: 7, (1999), pp. 639-644 Bibliographic Page Full text |
| [7]. | Caffrey C.R., Scory S., Steverding D., "Cysteine proteinase of trypanosome parasites: novel targets for chemotherapy", Curr. Drug Targets, Volume: 1, (2000), pp. 155-162 |
| [8]. | Caffrey C.R., Hansell E., Lucas K.D., et al. "Active site mapping, biochemical properties and subcellular localization of rhodesain, the major cysteine protease of Trypanosoma brucei rhodesiense", Mol. Biochem. Parasitol., Volume: 118, (2001), pp. 61-73 Bibliographic Page Full text |
| [9]. | Lonsdale-Eccles J.D., Grab D.J., "Lysosomal and non-lysosomal peptidyl hydrolases of the bloodstream forms of Trypanosoma brucei brucei", Eur. J. Biochem., Volume: 169, (1987), pp. 467-475 |
| [10]. | Authié E., "Trypanosomiasis and trypanotolerance in cattle: a role for congopain", Parasitol. Today, Volume: 10, (1994), pp. 360-364 |
| [11]. | Troeberg L., Pike R.N., Morty R.E., Berry R.K., Coetzer T.H.T., Lonsdale-Eccles J.D., "Proteases from Trypanosoma brucei brucei. Purification, characterisation and interactions with host regulatory molecules", Eur. J. Biochem., Volume: 238, (1996), pp. 728-736 |
| [12]. | Scory S., Caffrey C.R., Stierhof Y.-D., Ruppel A., Steverding D., " Trypanosoma brucei: killing of bloodstream forms in vitro and in vivo by the cysteine proteinase inhibitor Z-Phe-Ala-CHN2", Exp. Parasitol., Volume: 91, (1999), pp. 327-333 Bibliographic Page Full text |
| [13]. | Troeberg L., Morty R.E., Pike R.N., et al. "Cysteine proteinase inhibitors kill cultured bloodstream forms of Trypanosoma brucei brucei", Exp. Parasitol., Volume: 91, (1999), pp. 349-355 Bibliographic Page Full text |
| [14]. | Eakin A.E., Mills A.A., Harth G., McKerrow J.H., Craik C.S., "The sequence, organization, and expression of the major cysteine protease (cruzain) from Trypanosoma cruzi", J. Biol. Chem., Volume: 267, (1992), pp. 7411-7420 |
| [15]. | Hirumi H., Hirumi K., Doyle J.J., Cross G.A.M., "In vitro cloning of animal-infective bloodstream forms of Trypanosoma brucei", Parasitology, Volume: 80, (1980), pp. 371-382 |
| [16]. | Baltz T., Baltz D., Giroud C., Crockett J., "Cultivation in a semi-defined medium of animal infective forms of Trypanosoma brucei, T. equiperdum, T. evansi, T. rhodesiense and T. gambiense", EMBO J., Volume: 4, (1985), pp. 1273-1277 |
| [17]. | Merschjohann K., Sporer F., Steverding D., Wink M., "In vitro effect of alkaloids on bloodstream forms of Trypanosoma brucei and T. congolense", Planta Med., Volume: 67, (2001), pp. 623-627 |
| [18]. | Huber W., Koella J.C., "A comparison of three methods of estimating EC50 in studies of drug resistance of malaria parasites", Acta Trop., Volume: 55, (1993), pp. 257-261 Bibliographic Page Full text |
| [19]. | Rodriguez J., Deinhardt F., "Preparation of a semipermanent mounting medium for fluorescent antibody studies", Virology, Volume: 12, (1960), pp. 316-317 |
| [20]. | Woo J.-T., Sigeizumi S., Yamaguchi K., et al. "Peptidyl aldehyde derivatives as potent and selective inhibitors of cathepsin L", Biorg. Med. Chem. Lett., Volume: 5, (1995), pp. 1501-1504 Bibliographic Page Full text |
| [21]. | Shaw E., Mohanty S., Colic A., Stoka V., Turk V., "The affinity-labelling of cathepsin S with peptidyl diazomethyl ketones. Comparison with the inhibition of cathepsin L and calpain", FEBS Lett., Volume: 334, (1993), pp. 340-342 Bibliographic Page Full text |
| [22]. | Yasuma T., Oi S., Choh N., et al. "Synthesis of peptide aldehyde derivatives as selective inhibitors of human cathepsin L and their inhibitory effect on bone resorption", J. Med. Chem., Volume: 41, (1998), pp. 4301-4308 Bibliographic Page |
| [23]. | Grab D.J., Wells C.W., Shaw M.K., Webster P., Russo D.C.W., "Endocytosed transferrin in African trypanosomes is delivered to lysosomes and may not be recycled", Eur. J. Cell Biol., Volume: 59, (1992), pp. 398-404 |
| [24]. | Steverding D., Stierhof Y.-D., Fuchs H., Tauber R., Overath P., "Transferrin-binding protein complex is the receptor for transferrin uptake in Trypanosoma brucei", J. Cell. Biol., Volume: 131, (1995), pp. 1173-1182 |
| [25]. | Kabiri M., Steverding D., " Trypanosoma evansi: demonstration of a transferrin receptor derived from expression site-associated genes 6 and 7", J. Parasitol., Volume: 87, (2001), pp. 1189-1191 |
| [26]. | Robertson C.D., North M.J., Lockwood B.C., Coombs G.H., "Analysis of the proteinase of Trypanosoma brucei", J. Gen. Microbiol., Volume: 136, (1990), pp. 921-925 |
| [27]. | Schechter I., Berger A., "On the size of the active site in proteases. I. Papain", Biochem. Biophys. Res. Commun., Volume: 27, (1967), pp. 157-162 |
| [28]. | Caffrey C.R., Schanz M., Nkemgu-Njinkeng J., et al. "Screening of acyl hydrazide proteinase inhibitors for antiparasitic activity against Trypanosoma brucei", Int. J. Antimicrob. Agents, Volume: 19, (2002), pp. 227-231 Bibliographic Page Full text |
| [29]. | Hanzlik R.P., Thompson S.A., "Vinylogous amino acid esters: a new class of inactivators for thiol proteases", J. Med. Chem., Volume: 27, (1984), pp. 711-712 |
| [30]. | Palmer J.T., Rasnick D., Klaus J.L., Brömme D., "Vinyl sulfones as mechanism-based cysteine protease inhibitors", J. Med. Chem., Volume: 38, (1995), pp. 3193-3196 |
| [31]. | Engel J.C., Doyle P.S., Hsieh I., McKerrow J.H., "Cysteine protease inhibitors cure an experimental Trypanosoma cruzi infection", J. Exp. Med., Volume: 188, (1998), pp. 725-734 |
| International Journal of Antimicrobial Agents | ||
| Vol: 22 Issue: 2, August, 2003 | Article Full Text PDF (208 KB) | |
| pp: 155 - 159 | Bibliographic Page | |
| Full Text Article HTML | ||
| Get citation export (Reference Format) | ||
| |||||||||||||||||||||||||||||||||||||||
| Copyright © 2002 ScienceServer
LLC |
