|
||||||||||||||
|
Autonomous promotion pathway |
||||||||||
|
Arabidopsis fca mutants flower late in both long days and short days. This lateness can be suppressed by either a vernalization treatment or by growth in low red/far red light conditions. These phenotypic properties are shared by a sub-set of other late flowering mutants; namely, ld, fld, fca, fpa, fve and fy. These late-flowering recessive mutants have therefore been classified together, on the basis of these shared phenotypes and their epistatic interactions with other flowering time mutants, in a genetically defined group known as the autonomous pathway. The level of FLC mRNA is increased in these mutants, indicating that the autonomous pathway normally functions to prevent FLC mRNA accumulation.
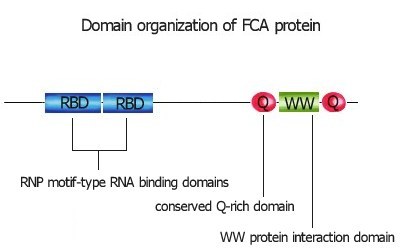
The gene we have been studying in this pathway is FCA (Macknight ref link). We map-based cloned FCA and found it encodes a protein with two RNP motif – type RNA binding domains and a protein-protein interaction domain known as the WW domain (EMBL Z82992, Z82989). This suggests that FCA acts in a ribonucleoprotein complex to regulate the floral transition.
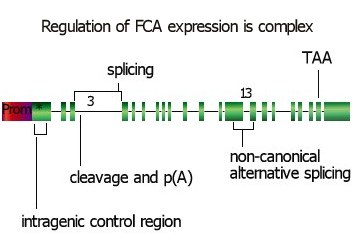
The control of FCA expression is (very unexpectedly) complex. The pre-mRNA is alternatively spliced, resulting in the production of at least four mRNAs, only one of which encodes full-length FCA. A major control point appears to be at intron 3: we can detect mRNAs that have either been cleaved and polyadenylated within intron 3 (which would be predicted to encode truncated FCA protein) as well as transcripts from which intron 3 has been correctly excised. A second alternative splicing event at intron 13 results in the production of a transcript that would encode an isoform of FCA that lacks the WW domain. The mechanism of this processing appears to be novel as the cis-elements involved are distinct from those required in conventional U2 or U12- dependent spliceosomal mediated pre-mRNA splicing. In addition to this control at the pre-mRNA level, we have recently found that the translation of FCA has unexpected properties and that intragenic sequences significantly influence FCA expression pattern. We have identified FY as an interacting partner with FCA through affinity purification, for western analysis and candidate gene approaches.
FY encodes a homologue of a well conserved eukaryotic protein that functions as a polyadenylation factor in yeast. Our current model is that FCA controls 3' end formation of specific transcripts by recruiting them to the polyadenylation / cleavage complex. The FCA / FY interaction regulates processing of the FCA transcript itself . Whether they regulate the FLC transcript directly is being explored.
Current areas of activity: 1) The controls of FCA expression are being dissected and their significance to the timing of the floral transition determined. 2) The hierarchy of flowering time control is being investigated by combining a transgene overexpressing FCA with different flowering time mutants - in the autonomous pathway, the FRI repression pathway and the photoperiod and vernalization promotion pathways. In addition we have isolated and characterised suppressor mutants of fca-1 (acf mutants). 3) We are undertaking experiments to address the mechanism by which FCA controls flowering time and are identifying the RNAs and proteins with which FCA interacts. 4) Inducible FCA expression systems are being generated. These will enable the timing of FCA function to be assessed and the molecular targets to be identified. 5) Genes required for FCA/FY down-regulation of FLC expression are being identified using a genetic analysis. 6) We are exploring the role of small/micro RNAs in FCA/FY function. 7) We are identifying the RNAs to which FCA
binds. |
||||||||||