Folding, Misfolding, and Lessons from a WW Domain
By Jason Socrates
Bardi
Protein misfolding can be such a difficult and abstract
subject that it becomes easy to lose sight of the simple
principles that scientists would like to understand regarding
how and why proteins misfold.
One reason can be illustrated by the discovery a doctor
made almost 100 years ago. When Dr. Alois Alzheimer examined a
post-mortem patient who died with an unusual mental illness in
1906, he found clumps of "amyloid" protein plaques in her
brain. These plaques are still a clear sign of the disease
that bears his namea disease now believed to inflict some 4
million Americans.
Many diseases are caused by the formation of protein
plaques inside the body. Amyloid-forming diseases like
Alzheimer's and Parkinson's are well known, but others include
a collection of over 80 rare amyloid diseases caused by the
misfolding of the protein transthyretin (TTR), which the liver
secretes into the bloodstream to carry thyroid hormone and
vitamin A. In diseases like familial amyloid polyneuropathy
(FAP), hundreds of these proteins misfold into structures
leading to microscopic fibril plaques, which deposit in
internal organs and interfere with normal function, sometimes
lethally.
Another reason for scientists' interest in protein
misfolding can be illustrated by a problem recently facing a
graduate student. The student was toiling away at a routine
laboratory procedure of expressing and purifying a small
soluble protein in bacteria, but every time the student
induced the expression of the protein, the protein molecules
would clump together on the nanoscale, form insoluble
"inclusion bodies" on the microscale, and crash out of
solution like a fried egg at the bottom of the test tube on
the macroscale.
In fact, understanding protein misfolding would have many
applications in basic biochemistry, where it could be used to
prevent such aggregation and advance basic laboratory methods.
As such, says Jeffery W. Kelly, who is the Lita Annenberg
Hazen Professor of Chemistry at The Scripps Research Institute
(TSRI) and vice president of academic affairs at TSRI,
understanding how and why proteins misfold is a high priority
for many scientists.
And, he adds, "In order to completely understand
misfolding, we have to understand how proper protein folding
takes place."
A Model For Beta Sheet Formation
Now two studies in an upcoming issue of Proceedings of
the National Academy of Sciences (PNAS) provide insight
into the folding process of what are known as beta sheet
structures. This is a common fold or "motif" wherein fully
extended peptide strands hydrogen bond to each other to form a
sheetthe same way that plastic teeth in a flat comb line up.
In one article, Kelly, his colleagues at The Skaggs
Institute for Chemical Biology at TSRI, and a group at the
University of Illinois examined the kinetics of beta sheet
formation using a protein fragment called the formin-binding
protein (FBP) WW domain. The "WW" refers to the fact that
there are two conserved tryptophan residues in the hundreds of
sequences comprising this domain family (tryptophan is
abbreviated "W").
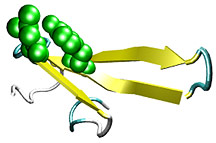
This FBP WW domain is a small protein fragment of a few
dozen residues that folds rapidly into its structure in tens
of microseconds. The structure is very basicits chain folds
to form three strands of a beta sheet connected by two loops.
Significantly, this structure is incredibly tolerant to
mutagenesistwo sequence-dissimilar WW domains from divergent
species will nevertheless fold into the same three-dimensional
structure. This tolerance is important to be able to mutate
residues of this protein and gauge which ones may be
particularly important for the folding of these WW domains.
And that's exactly what Kelly and his colleagues did. They
made changes to the protein, including mutating certain
residues, cutting off the end of the FBP WW domains, and
observing protein folding under temperature variations. They
observed this by monitoring a change in fluorescence as the
proteins adopts or changes its structure using a laser heating
and rapid fluorescence measurement technique employed by the
University of Illinois group.
What the researchers found was that temperature, mutation,
and truncation changes all have the ability to alter the
kinetics of the protein foldingchanging the pathway by which
these proteins fold, not their final structure.
The folding kinetics or the way that FBP and other WW
domain proteins fold can generally be divided into two
separate classesthe two-state folders and the three-state
folders.
Two-state folders form the characteristic beta sheet in
a single transition. That is, they go from one (unfolded)
state to a second (folded) state. Three-state folders, on the
other hand, go through an intermediate state, which means that
they have two transitions along their folding pathways
(unfolded to intermediate and intermediate to folded).
Kelly and his colleagues found that when the FBP WW domains
folded with three-state kinetics, the formation of a
particular loop between two individual beta strands is the
rate-limiting step for the folding of the beta sheet, meaning
that these connections form more slowly. They also found that
it is possible to "tune" the way the proteins folded by
changing the sequence or the temperature of the protein.
Specifically, Kelly and his colleagues were able to switch a
slow folding, two-transition WW domain into a more rapid,
single transition WW domain.
Kelly explains that higher temperatures, the removal of the
C-terminus of the protein, or cetain internal mutations
destabilize the intermediate state, so the proteins fold in a
single transition.
A Model of the Folding
A second PNAS paper, also scheduled for publication
soon, by TSRI graduate student John Karanicolas and Charles L.
Brooks III, professor of molecular biology at TSRI, predict
this behavior with a new model.
Karanicolas and Brooks model the folding kinetics of two
different WW domainswhich both fold into the classic WW beta
sheet, but via the two different mechanisms. Significantly,
Karanicolas and Brooks found through their simulations that
the origin of the three-state folding is a particular set of
residues that form mismatched contacts with each other in the
intermediate state.
These contacts occur during the formation of a particular
loop that must form before the beta strands can line up to
form a beta sheet. This process has the propensity to form a
misaligned loop in which the incorrect neighbors line up,
which is sort of like mismatching the buttons on your shirt.
This mismatched intermediate, Karanicolas and Brooks found,
is what drives the rate-limiting step. They observed this
misaligned loop and inferred that one particular residue that
is responsible for fixing the alignment is removed, the slow
phase gets slower.
Karanicolas and Brooks shared this result with Kelly, and
Kelly and his colleagues made mutations to the proteins,
tested their folding, and verified the prediction
experimentally.
What emerges from both of these studies is a complicated
description best explained by a simple analogy: the folding of
the WW domain is something like buttoning up your shirt in the
morning. If you misalign the buttons, it takes time to
unbutton the shirt and button it up again.
In other words, it's faster to button your shirt right the
first time than to button it incorrectly and have to fix it,
and it's possible to "tune" the process by adjusting
surrounding conditions, like the light in the room. Turn up
the dimmer, and you can see the shirt better. The better you
see, the less likely you are to make a mistake buttoning your
shirt, and the faster you will be able to do it.
"This [research] was an extraordinarily good set of
interactions," says Brooks. "Jeff [Kelly] and his colleagues
were really listening to our theoretical input, and we were
getting all this wonderful feedback that helped us understand
our models better and led, ultimately, to the correct picture
of the folding for this protein."
To read the article "Tuning the free energy landscape of a
WW domain by temperature, mutation, and truncation" by Houbi
Nguyen, Marcus Jager, Alessandro Moretto, Martin Gruebele, and
Jeffery W. Kelly, please see: http://www.pnas.org/cgi/doi/10.1073/pnas.0538054100.
To read the article "The structural basis for biphasic
kinetics in the folding of the formin-binding protein WW
domain: Lessons for protein design?" by John Karanicolas and
Charles L. Brooks III, please see: http://www.pnas.org/cgi/doi/10.1073/pnas.0731771100.

|