[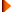 Scroll to first occurrence on
this page] Scroll to first occurrence on
this page] |
| [Scroll to first occurrence on
this page] |
All of the aceticlastic methanogens belong to this family, as well as all methanogens that disproportionate methanol, methylamines, or other methyl-containing compounds. The only other species that catabolize methanol are Methanosphaera species, which require H2 to reduce it to methane. The family Methanosarcinaceae can be divided into three physiological and morphological types: 1) the genus Methanosarcina, which contains coccoid and pseudosarcinal cells and can disproportionate methanol and catabolize acetate and H2+CO2; 2) the genus Methanosaeta, which grows only by the aceticlastic reaction; 3) the halophilic, methylotrophic organisms of the genera Methanolobus, Methanococcoides, and Methanohalophilus. Phylogenetically, only the first of these groups is composed of closely related species (Fig. 4).
Methanosarcina species disproportionate methanol and methylamines, forming CH4 and CO2 in a ratio of approximately 3:1 (Table 14). Most strains also use acetate, and most also use H2 to reduce either CO2 or methanol to CH4. Formate is never used. Cells appear to have a protein cell wall adjacent to the cytoplasmic membrane. External to the protein wall, many strains have a layer of methanochondroitin, referred to as a matrix. Cells without the matrix are individual, irregular cocci, but when the matrix is present, packets or pseudoparenchyma are formed (Fig. 7). Within the packets, the cells are irregular in shape, the division planes are not always perpendicular, and the volume of the daughter cells is not always equal. Three morphotypes of Methanosarcina have been described. Morphotype 1 includes some strains of M. barkeri that form aggregates of packets of cocci termed pseudoparenchyma, which may be visible to the unaided eye (Zhilina, 1976). Morphotype 2 includes M. vacuolata, which forms small packets and tends to remain dispersed and settles slowly in liquid medium. Morphotype 3 includes M. mazei, which forms single cocci, packets of pseudococci, and large macrocysts (Zhilina and Zavarzin, 1979b; Robinson, 1986).
Fig. 7. Methanosarcina barkeri strain MS. Bar = 10 ¼m. (Courtesy of D. Boone.)


While morphology and substrate range are important characteristics of Methanosarcina, they are not always reliable indicators of species differences. These characteristics frequently vary when closely related strains are tested, or the same strain may give varied results under different conditions. For example, a single enzyme may be responsible for changing large pseudoparenchymal aggregates into individual coccoid cells, and this enzyme may be produced only under certain growth conditions (Harris, 1987; Sowers and Gunsalus, 1988; Xun et al., 1988). The ability to use substrates often depends on the conditions of the inoculum (Boone et al., 1987), and the ability to form gas vacuoles is not a constant characteristic of strains (R. A. Mah, personal communication). Thus, variations in these characteristics within species may be commonplace, and they may not always be reliable for the placement of strains into species.
The diameter of the cocci or cells within the aggregates are generally 1-2 ¼m, although both smaller and larger cells are observed. The cells of Methanosarcina are nonmotile, and in those species that stain Gram negative, the cells lyse in the presence of sodium dodecyl sulfate. Cells of M. vacuolata contain numerous gas vacuoles tightly clustered in the cytoplasm. The Methanosarcina do not require additional carbon sources, and species that use H2 + CO2 as a substrate for methanogenesis can grow autotrophically. However, peptones or yeast extract are stimulatory for M. mazei, M. vacuolata, and M. frisia, and riboflavin is stimulatory or required for at least one strain of M. barkeri (Scherer and Sahm, 1981a). The chemicals 4-aminobenzoate and calcium chloride (0.7 mM) are required for growth of M. thermophila. M. barkeri grows with dinitrogen as its sole nitrogen source (Bomar et al., 1985; Murray and Zinder, 1984). Methionine, cysteine, thiosulfate, and elemental sulfur are also sulfur sources for some strains of M. barkeri (Mazumder et al., 1986; Scherer and Sahm, 1981b).
M. barkeri, M. mazei, M. thermophila, and M. vacuolata were isolated from anaerobic digestors. M. barkeri is also found in freshwater and marine sediments, rumens of ungulates, and animal-waste lagoons. M. mazei has been found in garden soil, sewage sludge, and various other sources. M. vacuolata is common in freshwater sediments, marshes and wetlands. M. acetivorans and M. frisia were isolated from marine sediments and for optimal growth require 0.2 and 0.3-0.4 M NaCl, respectively. M. acetivorans also requires 0.05-0.10 M magnesium for optimal growth. In addition to the type strain of M. thermophila, two strains of thermophilic Methanosarcina have been described (Ollivier et al., 1984; Touzel et al., 1985), whose nutritional characteristics resemble those of M. thermophila. However, both the pseudoparenchymal and coccoid morphologies are present, and the taxonomic status of these latter strains is uncertain.
Eight species and five genera of obligately methylotrophic, irregular cocci have been described (Table 15). In addition, two species of Methanolobus, M. siciliae and M. vulcani, have been proposed, but a complete description is unavailable (Stetter, 1989). At present, the taxonomy of these organisms is uncertain. Therefore, they have been arranged in the table in the order of increasing optimal salinity for growth, which varies from marine to extremely halophilic. Phylogenetic studies by RNA sequencing, RNA-DNA hybridization and DNA-DNA hybridization suggest that Methanococcoides methylutens, Methanolobus tindarius, Methanohalophilus zhilinae, M. mahii, and Methanohalobium evestigatum probably represent five different genera rather than only four (Sowers et al., 1984b; Chumakov et al., 1987; also see Fig. 4). The placement of the three remaining species listed in Table 15 is not known.
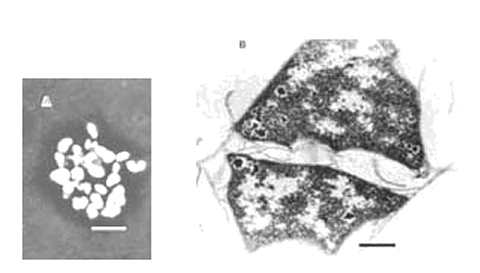
These species grow by disproportionation of methanol, methylamines, and other C-1 compounds. H2, formate, and acetate are not substrates. Cells are small irregular cocci, and some species form small aggregates. Cells of M. evestigatum are flat and polygonal, and they often occur in sheets (Fig. 8). Methanococcus halophilus produces slime. All the species are nonmotile, except for Methanolobus tindarius, which has monotrichous flagellation. With exception of Methanohalobium evestigatum, they stain Gram negative and lyse rapidly in dilute sodium dodecyl sulfate (or Sarkosyl for Methanococcus halophilus). These species are mesophilic, and their temperature optima are 30-35°C with three exceptions: the temperature optimum for Methanolobus tindarius is 25°C; the temperature optimum for Methanohalophilus zhilinae is 45°C; the temperature optimum for Methanohalobium evestigatum is 50°C. Methanohalophilus zhilinae and M. oregonense are also alkalophilic.
Fig. 8. Methanohalobium evestigatum. (A) Cells and aggregates, anoptral contrast Reichert. Bar = 5 ¼m. (B) Ultrathin section of late exponential phase cells. Layers of the envelope slip from the cell surface. Nuclear region and star-shaped granules (possibly glycogen) are present. Bar = 0.5 ¼m. (Courtesy of T. Zhilina.)
In addition to NaCl, M. euhalobius and M. mahii require high concentrations of magnesium and calcium for good growth. M. methylutens and M. oregonense require high concentrations of magnesium. Although the organic growth factors of the type strain of M. mahii are not known, strain SF-1, which appears to be closely related, requires yeast extract. Vitamins stimulate the growth of M. tindarius, and peptones that of M. zhilinae. In contrast, greater than 0.05% yeast extract or peptones inhibit the growth of M. halophilus and M. evestigatum.
M. tindarius and M. methylutens were isolated from marine sediments. M. mahii, Halomethanococcus doii, M. zhilinae, and M. oregonense were isolated from sediments of saline or saline and alkaline lakes. M. halophilus was isolated from a sample containing the cyanobacterial mat and associated mud of a stromatolite in Shark Bay, Australia. M. evestigatum was isolated from a cyanobacterial mat in a hypersaline lagoon close to Arabat, Sivash. M. euhalobius was isolated from the stratal liquid of an exploratory oil well.
In contrast to the other genera of the family Methanosarcinaceae, which are coccoid, the species of Methanosaeta (formerly called Methanothrix) are sheathed rods. These organisms grow only by the aceticlastic reaction (Table 16). Acetate is the only substrate for methanogenesis, and H2, formate, and methyl compounds are not utilized. Because of their very long generation time, 1 to 3 days under optimal conditions, pure cultures have been difficult to obtain. For instance, the original culture of "M. soehngenii" is not pure, and vancomycin is required to suppress the growth of contaminants (Patel and Sprott, 1990; Touzel et al., 1988). Likewise, the description of M. thermoacetophila is based upon a monoculture, although a pure culture has recently been obtained. Two other pure cultures have also been described. M. concilii appears to be closely related to the methanogen in "M. soehngenii" cultures, and strain CALS-1 appears to be similar to M. thermoacetophila, but insufficient cell mass has been obtained to complete the species description.
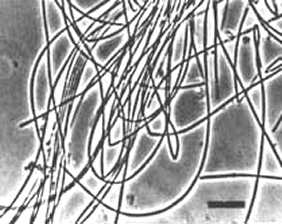
Methanosaeta is a short, fat, nonmotile rod with flat ends. Cells stain Gram negative and are sheathed. Except for CALS-1, stationary cultures may form long filaments with a contiguous sheath (Fig. 9). The filaments may form large bundles, mats, or flocs. In shaken cultures, the filaments tend to fragment into short filaments or individual cells. M. thermoacetophila and CALS-1 contain gas vacuoles and M. concilii produces a capsule.
Fig. 9. Methanosaeta concilii. Bar = 10 ¼m. (Courtesy of D. Boone.)
Marine or halophilic species have not been described, and NaCl does not need to be added to the media above trace amounts. The pH optima of "M. soehngenii", M. concilii, and CALS-1 are 7.4-7.8, 7.1-7.5, and 6.5, respectively. Yeast extract inhibits the growth of M. concilii, and penicillin inhibits the growth of "M. soehngenii" and M. thermoacetophila. CALS-1 is very sensitive to the reducing agents in the medium, and 1 mM sulfide and 1 mM 2-mercaptoethanesulfonate appear to be optimal.
Methanosaeta is widely distributed in nature. Its distinctive morphology has allowed it to be identified in sewage sludge, digestors, animal wastes, and sanitary landfills. Monocultures of the thermophiles have been obtained from different types of thermophilic digestors and thermal sediments. "M. soehngenii" and CALS-1 were obtained from sludge digestors. M. concilii was obtained from a pear waste digestor. M. thermoacetophila was obtained from the sediments of a thermal lake.